Az orr az egyik legfinomabb érzékszerv. De mi valójában a szaglás, és mennyire tudjuk megjósolni, hogy milyen vegyületek idéznek elõ különbözõ szagokat? teszi fel a kérdést Charles Sell, a Quest International szerves kémiai részlegének vezetõje. A Chemistry in Britain c. folyóiratban megjelent cikket kissé rövidítve közöljük.
Az állatvilágban a "kémiai érzékelés" szerepe messze felülmúlja a többi érzékelését, csak néhány madár és fõemlõs hagyatkozik inkább a látásra a szaglás vagy az ízlelés helyett. A kémiai jelek érzékelése lehetett az egyszerû szervezetek legelsõ érzékelési módja. Fontossága ellenére mégis keveset tudunk róla. A szagokat hasonló receptorok érzékelik, mint a hormonokat, és a látásban is hasonló receptorok mûködnek közre. A szaglás és az ízlelés folyamatának feltérképezése nemcsak tudományos, hanem kereskedelmi szempontból is jelentõs. Az illatszereket gyártó Quest International kutatócsoportjának vezetõjeként elsõsorban az foglalkoztat, hogyan jósolható meg az új molekulák illata.
A kölnik eleinte kizárólag természetes anyagokból készültek. A XIX. században azonban megindult a szerves szintézis, s a vegyipar új illatszereket állított elõ. Az elsõ mesterséges illatanyagok sokszor furcsa felfedézések nyomán születtek. Egy vegyész, A. Baur például robbanószerekkel kísérletezett 1888-ban, amikor felfedezte, hogy egyes nitro-benzol-származékok mósuszillatúak. Az elválasztás, a tisztítás és a szerkezetmeghatározás fejlõdésével sok természetes illat titkára fény derült, s megkezdõdhetett a hasonló anyagok gyártása. Míg a természetes olajok és kivonatok rendszerint több komponens keverékei, a szaganyagok általában jól definiált illattal jellemezhetõk.
Az illatanyagok tervezéséhez természetesen meg
kell értenünk a "mûködésüket". A szagérzékelés
modellezésére számos elmélet született.
A legfurcsább talán Arisztotelész sugárzási
elmélete, mely szerint az illatos anyagok sugárzást
bocsátanak ki, s a szaglószerv ezeket a sugarakat fogja fel.
W. K. Miles és L. H. Beck 1947-ben azzal az állítással
próbálta alátámasztani az elméletet,
hogy a legyek a méz illatát olyan edényen át
is megérzik, amely a távoli infravörös spektrumban
átlátszó. A kísérletet azonban sohasem
ismételték meg, és az elméletet ma már
nem veszik komolyan. Ugyancsak elvetették M. M. Mozell kromatográfiás
(1970), P. Gavaudan termodinamikai aktivációs (1948) és
J. T. Davies membránáthatolási elméletét
(1971). Érdekes módon ezek az elképzelések
a szagmolekula oldhatóságából és/vagy
illékonyságából indulnak ki, tehát olyan
tulajdonságokból, amelyek a molekula vándorlásában
játszanak szerpet, de nem vesznek feltétlenül részt
a jelek keltésében.
Felismerés kontra rezgés
Az elfogadhatónak tekintett elméleteket két csoportba sorolnám aszerint, hogy "felismerési" vagy "rezgési" teóriáról van-e szó. Mindkét csoport hipotézisei arra építenek, hogy a szagmolekula érintkezésbe lép egy receptorral, és ennek hatására a receptort taralmazó sejt idegimpulzust hoz létre. A felismerési elméletek szerint az érintkezés annak köszönhetõ, hogy egy fehérje (a receptor) felismeri a szubsztrátot (a szagmolekulát). A rezgési elméletek azt állítják, hogy a receptor valahogy érzékeli a szubsztrát egyik molekularezgését vagy a rezgések egy csoportját.
Az elsõ felismerési elmélet, Epikurosz elmélete, feltételezi, hogy az illatos anyagok apró részecskéket atomokat adnak le, és ezeket az orr érzékeli. Epikurpsz úgy gondolta, hogy a sima, kerek atomok édes, a hegyes atomok ingerlõ, savas illatot keltenek. A legismertebb modern felismerési elmélet John Amoore-tól származik. Amoore szerint az orrban korlátozott számú, különbözõ receptor van, ezek mindegyike adott molekulaalkot ismer fel, és amikor inger éri, egy alapszagnak megfelelõ jelet állít elõ. Az alapszagok hasonló célt szolgálnak, mint az alapszínek a színérzékelésben. Tehát a sokféle illat alapillatok keveréke, mint ahogy a színárnyalatok vörös, kék és sárga színbõl állnak össze. Amoore elõször úgy próbálta meghatározni ezeket az alapszagokat, hogy megkereste a szagok leírására szolgáló leggyakoribb szavakat. Ennek alapján a következõ hét alapszagot állapította meg: éteres, kámforos, mósuszos, virágillatú, mentolos, csípõs, rotható. Mindegyik fajta kémiai és sztérikus tulajdonságait tanulmányozta. Az elsõ öt esetben alak szerinti receptorokat tételezett fel, a csipõs szagot pozitív, a rothadót negatív töltésû részecskéknek tulajdonította.
Késõbb azt is feltételezte, hogy ha léteznek ezek az alapszagok, akkor meg kell felelniük a szagláshiány különbözõ eseteinek (a szagláshiányban szenvedõk egyes szagokat nem érzékelnek). A különbözõ szagláshiányokat és alapszagokat nagyszámú ember vizsgálatával próbálta megállapítani. A kísérletek alapján újabb alapszagokat vezetett be, mert péládul a négy legyakoribb szagláshiány közül (mósusz, szantálfa, ámbra és vizelet) csak az egyikhez tartozó szag szerepelt a kezdeti együttesben.
Két másik felismerési elmélet is figyelmet érdemel. M. G. J. Beets 1968-ban vetette fel, hogy a szagmolekulát a funkciós csoportja "állítja" a receptorhoz, és a szagot a receptor elé tárt "molekulaprofil" határozza meg. A másik egy moldáviai kutatócsoport elektrontopológiai hipotézise. Eszerint a felismerésben a szagmolekula külsõ elektronjai és a "szagháromszögek" játszanak szerepet.
A füst-, a hús-, az ámbra- és a mósuszillat esetében meghatározták, hogy az adott illattípushoz milyen atomi háromszögnek és milyen elektromos tulajdonságoknak kell kialakulni a molekulában. A háromszög mérete és alakja szagonkényt változó, de rendszerint egy oxigén- és két hidrogénatomból áll. A mósuszillathoz szükség van egy másik molekularészre is, amely két, egymással adott geometriai kapcsolatban álló hidrogénatomot tartalmaz. A csoport 1966-ban finomította az ámbrára vonatkozó szabályt: két darab, az eredetinél kissé nagyobb molekularészre van szükség, amelyekben a külsõ elektronok helyett az atomi töltés határozza meg a szükséges elektromos tulajdonságokat.
Az elsõ rezgési elméletet G. M. Dyson fogalmazta meg 1937-ben. Dyson úgy gondolta, hogy az orrbeli receptor a szagmolekulák rezgését érzékeli, s az agy ezeket értelmezi szagként. Az elképzelést Robert Wright fejlesztette tovább: az 1960-as és 70-es években korrelációt talált az infravörös spektrum és a szag között. Dyson elméletnének gyengéje, hogy a rezgés érzékelésére nem ad választ.
Luca Turin feltételezi, hogy a receptorfehérjében,
amelynek a NADPH és az cinkionok az "elektródjai" elektromos
potenciálgát van. Az elektronok csak akkor juthatnak át
a gáton, ha az elektródok közé szagmolekula kerül.
Az elektronok az alagúthatás révén jutnak át
a szagmolekula elektronpályáin, s leadott energiájuk
rezgéseket gerjeszt a molekulában.
Minden az agyban dõl el
Ezek az elméletek alig-alig épülnek biológiai információkra. A teljes folyamat megfigyelésén alapulnak, pedig az illat képzõdése és agybeli felismerése között számos lépés van.
Az agy a szagokat a jellegük, az intenzitásuk és a tartósságuk szerint ismeri fel. Mindhárom tulajdonság szubjektív. A legtöbb vizsgálatot a jelleggel végezték, mert legalábbis felszínesen ez mérhetõ a legkönnyebben. A szag leírása azért nehéz, mert nincsenek rögzített viszonyítási pontok. A korrelációkeresés nehézsége rendszerint abban rejlik, hogy nem megfelelõ szagosztályozó rendszereket használunk. Kísérleteink során például kiderült, hogy a "gyümölcsillat" kategórája zavart okozhat, hiszen másfajta molekula idézi elõ az alma, és megint más a körte illatát. Márpedig a szerkezetszag korreláció vizsgálatához értelmes paramétereket kell használnunk.
A szagelméletek gyakori hibája, hogy nem tesznek különbséget az ok és az okozat között. Sokszor mutatunk ki korrelációt és oksági kapcsolatot két paraméter között anélkül, hogy megkérdeznénk: nem a két jelenség közös okát neveztük-e korrelációnak. Ilyen például az a feltevés, hogy adott szagmolekulák infravörös spektruma és szaga közötti korreláció azt bizonyítja, hogy a szagot bizonyos rezgések keltik. Valójában a szag is, a spektrum is közös okra, a molekulaszerkezetre vezethetõ vissza.
A szerkezetszag korreláció a szag "mérésén" alapszik. De a szag nem fizikai tulajdonság. Csak az agyban létezik; nem tételezhetjük fel, hogy a receptorban is létezik. Semmi sem bizonyítja, hogy a receptorok egyes szagokra vagy szagítpusokra lennének hangolva. Igen valószínû, hogy a felismerési folyamat a szagmolekulák fizikai és kémiai tulajdonságain alapszik, és szag csak akkor keletkezik, amikor az agy értelemzi a szaglóidegek jeleit.
A szerkezetszag korreláció egyik híres problémája a keserûmandula illata. A leginkább a benzaldehid és a hidrogén-cianid idézi elõ ezt az illatot. A benzaldehidhez hasonló és a benzaldehidet helyettesítõ vegyületek vizsgálata azt mutatja, hogy minél inkább hasonlít egy molekula tér- és elektronszerkezete a benzaldehidéhoz, annál inkább megközelíti a mandula illatát. A hidrogén-cianid persze nagyon különbözik a benzaldehidtõl. A mandulareceptor feltételezésével nagyon nehezen magyarázhatjuk meg az eredményeket. De ha arra gondolunk, hogy a mandulaillat csak az agyunkban létezik az idegimpulzusok eredményeként, már nem vagyunk bajban. Hiszen feltehetjük, hogy az agy megtanulta a kétféle jelsorozat (a benzaldehiddel és a hidrogén-cianiddal keltett jelek) hasonló értelmezését, mert a természetes mandulában a két molekula együtt jelenik meg az íz elõanyaga, az amigdalin (a benzaldehid-cianohidrin glikozidja) lebontásának eredményeként.
Nagyon sok mindent nem tudunk még a szaglásról
és az illatok megkülönböztetésérõl.
A kötõfehérjék, a hám "földrajzi"
viszonyai, az idegi impulzusok haladási útvonalai mind szerepet
játszhatnak a felismerési folyamatban. Azok a hibák,
amelyeket a szaglási folyamat modellezésekor elkövetünk,
arra figyelmeztetnek, hogy nem mondhatunk le a logikus gondolkodásról,
meg kell kérdõjeleznünk a feltevéseket, menekülnünk
kell a paradigmáktól különösen akkor, ha
nagy és bonyolult problémákat kell megoldanunk.
Irodalom
J. E. Amoore, Molecular basis of odour. Springfield, Illinois, US :
Charles C. Thomas, 1970.
K. Axel, Sci. Amer., p 130, October 1995.
H. Breer, K. Raming and J. Krieger, Biochem. Biophys. Acta, 1994, 1224,
277.
H. Kandori, Chem. Ind., 1995, 18, 735.
P. Laffort in Relationships between molecular structure and olfactory
activity, in odours and deodorization in the
environment, G. Martin and P. Laffort (eds). Weinheim: VCH, 1994.
K. J. Rossiter, Chem. Rev., 1996, 96 (8), 3201.
L. Turin, Chemical Senses, 1996, 21 (6), 773.
R. H. Wright, The sense of smell, Baton Rouge, Florida, US: CRC Press,
1982.
I. B. Bersuker and A. S. Dimogol in Reviews in computational chemistry,
K. Lipkowitz and D. Boyd (eds), p 423. New
York: VCH, 1991.
A szag biológiája
Az ember a szagot a szaglóhámon érzékeli, amely az orrüregben, a szemmel csaknem egy vonalban helyezkedik el. A hámon számos receptorsejt van. Ezek a sejtek idegimpulzusokat küldenek az agy szaglógumójába. Valószínû, hogy egy receptortípus minden jele ugyanabba a részbe jut a szaglógumóban.
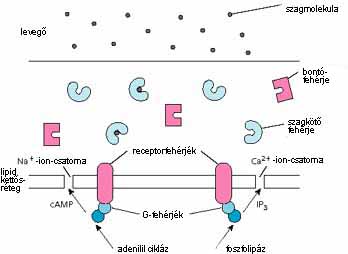
Az ábra egy receptorsejtet mutat. A szagmolekulák a levegõbõl az orr nyálkahártyájára kerülnek, a receptorsejt köré. Nem tudjuk, hogyan jutnak be a sejtbe, de mivel valamelyest oldódnak a vízben, létrejöhet a diffúzió bár ez nem magyarázza meg, hogy az vízben oldhatatlan szénhidrogéneknek, például az izooktánnak miért van szaga. Egy másik elképzelés szerint a szagmolekula szagkötõ fehérjék segítségével jut át a nyálkahártyán. A legelfogadottabb hipotézis azt mondja, hogy a szagkötõ fehérjék a szagmolekulát a sejtek felszínén ülõ receptorfehérjékhez szállítják, de az is lehet, hogy a kötõfehérjeszagmolekula komplex kötõdik a receptorfehérjéhez. A különbözõ szagkötõ fehérjék más-más affinitást mutatnak a szagmolekulákkal és a receptorfehérjékkel szemben. A nyálkahártyában bontófehérjék is vannak talán azért., hogy folyamatosan eltávolítsák a szagmolekulákat vagy szagmolekulakötõfehérje komplexeket, és így elõsegítsék, hogy a jelkeltés megszûnjön a szagforrás eltávolítása után.
A receptorfehérjék a sejt lipidmembránjában foglalnak helyet. A fehérje váza hétszer halad át a membránon, s a fehérje egyik vége a sejten a kívül, a másik a sejten belül van. A membránon áthaladó részek a-hélixek. Ezek az a-hélixek együtt hengert formálnak. A feltevések szerint ebben vannak a szagmolekulák kötõhelyei. Két kutatócsoport tanulmányozta a patkány OR5 nevû szagreceptor-fehérjéjének lehetséges kötõhelyeit. Az egyik csoport a 2-metil-3-(4-tetrabutil-fenil)-propanal, a másik a 3,7-dimetil-4-hidroxi-oktanal lehetséges kötõhelyét találta meg. A két kötõhely a fehérje hasonló területein van, s mindkét vegyület illata a gyöngyvirágéra emlékeztet.
A szagmolekula és a receptorfehérje kölcsönhatása megváltoztatja a fehérje szerkezetét, és a receptorhoz tartozó G-fehérje vagy a foszfolipáz C-t, vagy az adenilil-ciklázt aktiválja, s ennek nyomán másodlagos hírvivõ, vagy inozitol-trifoszfát (IP3), vagy ciklikus adenozin-trifoszfát (cAMP) kerül a sejtbe. Ezeknek a másodlagos hírvivõknek a hatására a sejtmembránban kinyílnak a kalcium- és nátriumion-csatornák, és idegimpulzusok képzõdnek. A másodlagos hírvivõk foszforilezéssel elindítják a receptor dezaktiválását. Az újbóli aktiváláshoz defoszforilezés szükséges.
A receptorfehérjéket kódoló géncsalád
a legnagyobb genom, akár több ezer különbözõ
receptorfehérjét tartalmazhat. Egyetlen fajban azonban nem
jelenik meg minden receptorfehérje, s a meglevõk sem egyenletesen
oszlanak el a hámban. Ezért a hám különbözõ
területérõl érkezõ szagok intenzitása
is hozzájárulhat a szag azonosításához.