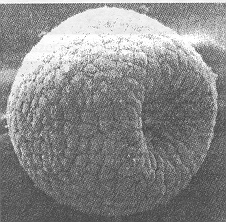
1. ábra. Tengerisün embrió a 10. sejtosztódás után. Az invaginációval együttjáró felületi alakváltozás első- vagy másodfajú fázisátalakulásként értelmezhető
Fizikai Szemle honlap |
Tartalomjegyzék |
Fizikai Szemle 1996/3 95.o.
Forgács Gábor
KFKI SZFKI és
Clarkson University, Potsdam, N.Y. USA
C.H. Waddington [1] az evolúcióelmélet neves szaktekintélye a "The Nature of Life" című könyvében írja. "A fejlődés egy többé-kevésbé gömbalakú tojásból indul. Ebből lesz egy élőlény, amely minden, csak nem gömb... Ezt nem képes megmagyarázni semmiféle olyan elmélet, amely pusztán kémiai kijelentésekre korlátozódik, mint például az, hogy gének szintetizálnak adott fehérjéket. Valahogyan be kell hozzuk a sztoriba azokat a fizikai erőket, melyek ahhoz szükségesek, hogy a fejlődés során az anyag a megfelelő helyre kerüljön és abból a helyes formák alakuljanak ki. " Waddington a fenti kijelentést több mint harminc évvel ezelőtt tette, de még ma sem mondható el, hogy a biológusok többsége maradéktalanul egyetértene vele:
Jelenleg két jól különválasztható nézet figyelhető meg a biológiában általánosan és különösképpen a fejlődéstanban. Az egyik nézetet D'Arcy Thompson [2] követői képviselik, akik főleg modellgyártó elméleti fizikusok és matematikusok. Eszerint (végletesen fogalmazva) a biológiai fejlődés minden szakasza megmagyarázható a fizika és a matematika mai állása alapján. A másik nézet szerint (melyet leginkább gyakorlati biológusok reprezentálnak), ismét végletesen fogalmazva, a fejlődés minden egyes mozzanatát a gének és a bennük rögzített utasítások determinálják. A fizikai folyamatok teljesen passzív alárendeltjei a gének által kiadott "parancsoknak". A cikk szerzője osztja a számukban folyamatosan növekvő azon kutatók véleményét, akik szerint egyik fenti megközelítés sem helyes: a biológiai jelenségek helyes értelmezése csak a genetikus és szomatikus (azaz fizikai) tulajdonságok kölcsönhatások figyelembe vételével kapható meg [3].
1. ábra. Tengerisün embrió a 10. sejtosztódás után. Az invaginációval együttjáró felületi alakváltozás első- vagy másodfajú fázisátalakulásként értelmezhető
Hogy ilyen kölcsönhatásnak kell léteznie, könnyű látni az alábbi példából. Az embrió a genetikus kód által definiált szabályok és a rendelkezésre álló "nyersanyag" alapján létrehozza a fejlődéséhez szükséges "építőanyagot". Ez jelentheti azokat a fehérjéket, melyeknek diffúzió, kemotaxis, differenciált adhézió stb. segítségével a fejlődő organizmus meghatározott pontjára kell eljutniuk (a kadherineknek például a sejt felületéhez). Ezt az anyagtranszportot a fizika törvényei szabályozzák: azok a mechanikai és kémiai változások, amelyek a transzport során végbemennek (lokális koncentrációváltozás, alakváltozás, adhéziós és kohéziós tulajdonságok megváltozása stb.), visszahatnak az "építőanyag-termelésre", vagyis a gének tevékenységére [4-6].
Az írás célja az, hogy néhány konkrét példán keresztül bemutassa, hogyan válhat a fizikus hasznossá az egyre fontosabb szerepet játszó biológiai kutatások számára.
Ma divatos az a mondás, hogy a fizikában nincsenek igazán izgalmas nyitott kérdések: mindent megoldottunk. Ez így valószínűleg túlzás, de azzal a ténnyel minden kezdő és fiatal fizikusnak tisztában kell lennie, hogy a teljesen elvont elméleti fizikai kutatásokra a világon mindenhol egyre kevesebb pénz áll rendelkezésre. A fizika a legrendszerezőbb diszciplína valamennyi természettudomány között, és mint ilyen, alkalmazása és alkalmazhatósága a biológiában is magától értetődő. A fizikus, vizsgálata során az adott rendszert annak legjellemzőbb tulajdonságainak megőrzésével igyekszik addig egyszerűsíteni, míg az általa ismert módszerekkel (Newton-egyenlet, Schrödinger-egyenlet stb.) kezelhető lesz. Az ilyen redukciós megközelítés óhatatlanul bizonyos specifikus tulajdonságok elhanyagolását eredményezi. Ezzel szemben a biológus tipikusan minden egyes rendszert szinte kizárólag annak specifikus tulajdonságain keresztül igyekszik megérteni. Az ilyen megközelítés kizárja az általánosítás lehetőségét. A biológiai rendszerek kutatása - bonyolultságuk miatt megköveteli a különböző szakterületen dolgozók együttműködését, és így a fizikus aktív részvételét is. A fizikus azonban csak akkor lesz hasznos tagja a csapatnak, ha a fentiekben elmondottak alapján megközelítési módszereiben hajlandó bizonyos változtatásokra.
Felületi jelenségek az embrionális fejlődésben
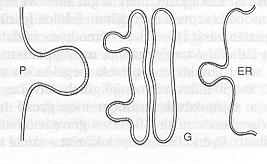
2. ábra. Felületi alakváltozások sematikus ábrázolása a sejt különböző membránszerű struktúráiban. (P = sejtfal, G = Golgi apparátus, ER = endoplazmás retikulum.)
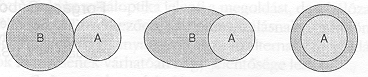
3. ábra. Különböző típusú egyedi sejtek, illetve sejtaggregátumok érintkezésekor végbemenő lehetséges folyamat sematikus bemutatása. A folyamat minden egyes fázisa egymással érintkező, de egymással nem elegyedő folyadékok tulajdonságaival értelmezhető.
Az 1. ábra egy tengerisün embriót mutat a tizedik sejtosztódás után. Az egyébként szinte tökéletes gömbalakú embrió egyik pontján ekkor egy betüremkedés (invagináció) jelenik meg. Kezdetét veszi a gasztruláció, vagyis az a folyamat, melynek során végülis kialakul a bélrendszer. Az invaginációt megelőző gömbalak egyetlen sejtrétegből áll, belseje üreges. A 2. ábrán sematikusan egyedi sejtek különböző membránstruktúráiban található kitüremkedések láthatók: a sejtfalon (P), a Golgi apparátusban (G), illetve az endoplazmás retikulumban (ER). Az 1. és 2. ábra közös vonása, hogy egy szinte tökéletes geometriai alakzatban (gömb vagy sík), lokálisan alakváltozás következik be. Az alakváltozás mindkét esetben réteges, hártyaszerű szerkezetben megy végbe: az 1. ábrán a réteget sejtek, míg a 2. ábrán lipoid molekulák alkotják. Az ilyen alakváltozások aktív statisztikus mechanikai kutatások tárgyát képezik [7]. Az alakváltozás vagy első- vagy másodfajú fázisátalakulás, mely a hártya felületi (görbületi) rugalmasságának, valamint a betüremkedést határoló görbe vonalfeszültségének versengéséből adódik [7].
A 3. ábra a phagocytózis vagy az egymással érintkező szövetek tipikus viselkedésének vázlatos bemutatására szolgálhat. Az első esetben A és B két különböző sejteket tartalmazó aggregátumot jelöl. Amikor ezek kapcsolatba kerülnek egymással, az egyik az ábrán látható módon "körülfolyja" a másikat. Az ilyen és ehhez hasonló folyamatokat Steinberg vizsgálta először szisztematikusan [8]. Steinberg feltételezte, hogy a bemutatott folyamat, mely az embrionális morfogenezis egyik leggyakoribb velejárója, megérthető az egymással nem elegyedő folyadékok elmélete alapján. Az általa bevezetett "differenciális adhézió hipotézis" szerint [8] a különböző szövetek viszkoelasztikus anyagok, melyek többek között jól definiált felületi feszültséggel jellemezhetők. Amikor kapcsolatba kerülnek egymással, egyensúlyi konfigurációban a nagyobb felületi feszültséggel rendelkező aggregátum (a 3. ábrán A-val jelölt) kerül belülre. (A phagocytózis hasonlóan magyarázható [9].)
Steinberg és munkatársai megpróbálták a szöveti aggregátumok felületi feszültségét megmérni és az eredményeket korrelációba hozni a különböző egymással érintkező sejthalmazoknál tapasztalt egyensúlyi konfigurációkkal [9]. A kapott eredmények meglehetősen pontatlanok voltak, mivel nem állt rendelkezésre alkalmas mérési eljárás. (A nagy viszkozitás miatt a felületi feszültségmérés ismert módjai, mint gyűrűs vagy csepp módszer, nem alkalmazhatók.) Steinberg és eme cikk írója együttműködésének eredményeként a közelmúltban sikerült olyan mérési eljárást kidolgozni, mely végül lehetővé tette számos embrionális szöveti aggregátum felületi feszültségének meghatározását [10, 11]. A berendezés vázlatosan a 4. ábrán látható. Az aggregátum, mely tipikusan 30-40 ezer élő sejtet tartalmaz, a mérést megelőzően minden esetben 200-350 mikron átmérőjű gömb. Mivel az aggregátumok a sajátjukéval majdnem megegyező fajsúlyú tenyészközegben vannak [10, 11], a gravitáció hatása elhanyagolható. Ilyen körülmények között a szinte tökéletes egyensúlyi gömbalak a véges felületi feszültség következménye. A mérés során az aggregátum két határoló felület közé van helyezve, majd ismert méretű kompressziónak van alávetve. Az összenyomott aggregátum alakjából, ismert módon (a Laplace-egyenlet segítségével. [12]) a felületi feszültség meghatározható. A méréseket eddig hat különböző szövetre végeztük el [11]. A kapott eredmények minden esetben összhangban voltak az aggregátum-párok tapasztalt egyensúlyi konfigurációival. Az 5. ábrán csirkeembrió szívsejtjeiből preparált aggregátumon végzett konkrét mérés látható.
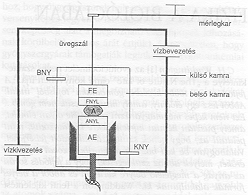
4. ábra. Embrionális szövetek felületi feszültségének mérésére szolgáló berendezés vázlata. A 30-40 ezer élő sejtet tartalmazó, 200-350 mikron átmérőjű aggregátum az alsó nyomólemezen (ANYL) nyugszik, mely a függőleges irányba mozgatható alsó egységhez (AE) van rögzítve. A felső nyomólemez (FNYL) a felső egységen (FE) keresztül, üvegszál segítségével egy elektromágneses mérleg karjához csatlakozik. A mérleg nullpozíciós elven működik, azaz az aggregátum összenyomásakor biztosítja, hogy FE ne mozduljon el. A nyomóerőt a nullpozíció fenntartásához szükséges mágneses tér, illetve ennek kialakítására szolgáló áram nagysága adja. Az aggregátum, AE és FE a belső kamrában helyezkednek el, mely 37 °C-on tartott szövetkultúra folyadékot tartalmaz. A belső kamra a befolyó nyíláson (BNY) keresztül tölthető fel, illetve az kifolyó nyíláson (KNY) keresztül üríthető ki. A belső kamrát egy külső kamra határolja. A külső kamrán keresztül áramoltatott víz biztosítja, hogy a szövetkultúra folyadék a fiziológiás körülményeknek megfelelő
37 °C-on legyen.

5. ábra. Egy a 4. ábrán ismertetett berendezéssel végzett konkrét mérés fényképfelvétele. a: 275 mikron átmérőjű embrionális csirke szívsejt aggregátum a kompressziót megelőző állapotban. b: Az a ábrán bemutatott aggregátum kompresszió alatt. A felső nyomólemezen kis légbuborék látható
Felmerülhet a kérdés, honnan tudjuk, hogy a fenti módon mért mennyiségek valóban kielégítik a felületi feszültség definícióját. A statisztikus mechanika szerint a felületi feszültség az egységnyi felület létrehozásához szükséges szabadenergia, és mint ilyen, az adott esetben függetlennek kell lennie mind az aggregátum méretétől, mind a kompresszió nagyságától. Méréseinket különböző méretű aggregátumokkal végeztük, azokat többszöri, fokozódó erősségű kompressziónak vetve alá. A hibahatáron belül minden esetben ugyanazt az eredményt kaptuk.
A fentiekben ismertetett berendezés alkalmas a viszkozitásnak, mint a folyadékfázist jellemző másik fontos fizikai paraméternek a meghatározására is. Az előzetes mérések azt mutatják, hogy az embrionális szövetek 104 Poise nagyságrendú viszkozitással rendelkeznek.
Az embrionális morfogenezis során a viszkoelasztikus anyagnak tekinthető megtermékenyített gömbalakú petesejtből alakváltozások sorozatának eredményeként az organizmus elnyeri végső alakját. Ahhoz hogy ezen alakváltozások a megfelelő pillanatban következzenek be, jól koordinált anyagáramlásra van szükség. A fejlődés eme kezdeti szakaszában az anyagáramlást leginkább felületi erők indukálják. Amennyiben a fenti gondolatmenet helyes, nyilvánvaló, hogy az embrionális szöveteknek, mint viszkoelasztikus anyagoknak a felületi feszültsége és viszkozitása meghatározó tényezője lehet az embrionális morfogenezis során végbemenő folyamatoknak. Ezen fizikai paraméterek ismerete lehetővé teszi, hogy konkrét jóslatokat tegyünk eme folyamatok időbeli menetére.
Mechanikai folyamatok szerepe biológiai jelátvitelben
Az élőlény nyitott rendszer. A környezetéből állandóan beérkező stimulusokat felfogja, továbbítja és végül fizikai vagy kémiai módon reagál rájuk. Ahhoz tehát hogy az organizmus normálisan funkcionálhasson, kifinomult jelzőrendszerrel kell bírnia. A ma elfogadottak alapján a szervezeten belüli jelek továbbítására három kémiai szignalizációs mechanizmust különböztetünk meg [13]. Az endokrin rendszer hormonokat használ, melyek a véredényrendszeren keresztül közlekedve érik el a target sejteket. A parakrin rendszer a szomszédos sejtek kommunikációját célozza, vagy lokálisan kiválasztott kémiai mediátorokon vagy a sejtközi állományon keresztül. A szinaptikus rendszert főleg az idegrendszer használja.
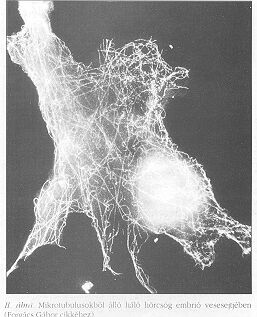
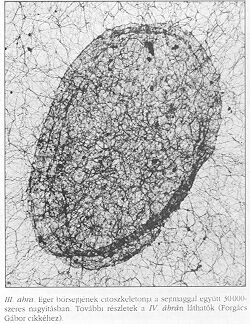
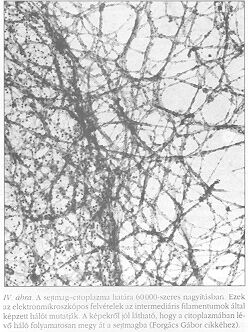
A továbbiakban a tárgyalást a parakrin rendszerre korlátozzuk, ahol a sejtek tipikusan receptor-ligandum párok segítségével jeleznek egymásnak. A parakrin jelátvitel első állomása a receptor-ligandum kötés kialakulása. A legtöbb sejtfelületi receptor egyben transzmembrán fehérje is, mely intracelluláris végződésén keresztül kapcsolódik a citoszkeletonhoz, a sejt belsejének architektúráját meghatározó vázhoz. A citoszkeletont aktin (I. ábra a hátsó borítón) mikrotubulus (II. ábra) és intermediáris filamentumok (III-IV. ábra) alkotják. Ezek a filamentumok láthatóan az egész sejtet behálózzák (beleértve a sejtmagot), és struktúrájukban rendezetlen pókhálóhoz hasonlíthatók.
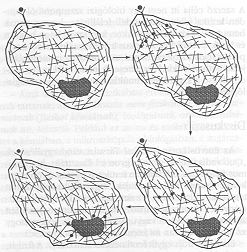
6. ábra. Az összefüggő perkolációs hálón alapuló mechanikai jelátvitel sematikus bemutatása.
Amikor a receptor-ligandum kötés kialakul, lokálisan energia szabadulhat fel, mivel a receptor fehérje alacsonyabb energiájú konfigurációba kerülhet. A receptor intracelluláris részének és a citoszkeletonnak a kapcsolatán keresztül ez az energiatöbblet az összefüggő filamentumrendszer mechanikai feszültségi állapotának megváltozását idézheti elő. Az eredeti kémiai jel ily módon elvben tisztán mechanikai úton, nevezetesen mechanikai deformáció formájában terjedhet a sejten belül, felhasználva a citoszkeletont. Mivel a filamentum-rendszer a sejtmagot is átszövi, a mechanikai jel, visszaalakulva kémiai energiává, elvben génaktivitást válthat ki (vagy felkapcsolhat vagy kikapcsolhat bizonyos géneket).
A fentiekben vázolt jelzési mechanizmus vázlata a 6. ábrán látható. A sejt felületén kialakuló jel tehát mechanikai úton, redundánsan (és így megbízhatóan) juthat el rendeltetési helyére. Mindezt az teszi lehetővé, hogy a citoszkeleton egy összefüggő hálót alkot. Az ilyen (rendezetlen) képződményeket a fizikában perkolációs hálók néven ismerjük; melyek tulajdonságait a statisztikus fizika módszereivel ma számos kutató vizsgálja [14]. Amennyiben a perkolációnak valóban szerepe lehet az intracelluláris biológiai jelek továbbításában, akkor eme hálók fizikai tulajdonságainak ismerete nagymértékben hozzájárulhat a biológia ma egyik legfontosabb és legizgalmasabb nyitott kérdésének megoldásához.
Hangsúlyozni kell, hogy az itt elmondottak egy szándékosan rendkívül leegyszerűsített modellt reprezentálnak. A fenti elgondolásokkal kapcsolatban az olvasóban számos kérdés merülhet fel (és kell is, hogy felmerüljön).
A szerző célja itt nem egy biológiai szempontból minden kritika felett álló modell felállítása, hanem annak bemutatása, konkrét példákon keresztül, hogy hogyan és hol kapcsolódhat a fizikus a biológiai kutatásokba. A perkolációs hálók esetleges szerepéről az intracelluláris jelátvitelben, sokkal részletesebb leírást talál az érdeklődő olvasó a szerző közelmúltban megjelent összefoglalójában [15].
Diszkusszió
Az Eötvös Loránd Fizikai Társulat vándorgyűlésének "Quo vadis fizika?" címe nagyon jól illusztrálja, hogy számos fizikus ma bizonytalan területen mozog és önmagát keresi. Vannak, akik azt állítják, hogy a fizika nagy nyitott témái vagy kimerültek (ez nem valószínű), vagy túl pénzigényesek (mint magfúzió). Mások szerint egyszerűen átmeneti időszakban vagyunk két robbanás között (hogy az utolsót pontosan mi jelenti, az nézőpont kérdése).
Eme cikk írója egyik fenti nézetet sem osztja. A fizikát, mint tudományágat nem lehet függetleníteni a társadalom szükségleteitől. Ez a kijelentés egyesek számára frázisként hangozhat, ez azonban a lényegen mit sem változtat. Természetesen mindig lesznek olyanok, akiknek majd megadatik - és meg is kell adni nekik -, hogy elvont problémákkal foglalkozhassanak, de amennyiben a jelen tendenciák megmaradnak, az ilyen kutatók száma tovább fog csökkenni. A csökkenés erőteljesen érezteti hatását az Egyesült Államokban (ahol a szerző jelenleg dolgozik). Egyetemi fizika tanszékeket szüntetnek meg vagy csoportosítanak át, ami pár évvel ezelőtt teljességgel elképzelhetetlen lett volna.
Mindezzel egyidejűleg azonban egy másik, sokkal inkább pozitív jelenség is megfigyelhető. Egyre több interdiszciplináris kutatócsoport alakult, illetve interdiszciplináris kutató intézet létesül, ahol előszeretettel alkalmaznak fizikust. Ezek túlnyomó része ma elsősorban biológiai jellegű vagy környezetvédelemmel kapcsolatos kutatásokra összpontosít. Itt jelentkezik a fentiekben említett társadalmi szükséglet.
Mit tehet ma egy kezdő vagy fiatal fizikus, akinek nagy valószínűséggel előbb vagy utóbb állásproblémával kell szembenéznie? Eleve otthagyja a fizikát és más pályát választ. Ignorálhatja az érvényesülő tendenciákat, és ha megvannak rá a lehetőségei, olyan kutatást folytat, ami neki a leginkább tetszik. Ha jelenleg esetleg léteznek is ezek a lehetőségek, az adott feltételek mellett azonban még semmiféle garancia nincs arra, hogy ezek a későbbiekben is fennmaradnak. Végül megpróbálhatja tudását és munkamódszereit egy interdiszciplináris csapat tagjaként hasznosítani. Ez természetesen további ismeretanyag elsajátítását igényli, mivel meg kell tanulnia azt a "nyelvezetet", mely szükséges a csapat tagjaival való kommunikációhoz. A jelen cikkben ismertetett és felvázolt biológiai-fizikai problémák eme utóbbi választási lehetőséghez kívántak segítséget, illetve támpontot nyújtani.
 |
 |
 |
 |
Irodalom